Emergent Complexity in Multi-Planetary Ecosystems
30 June 2017
Friday
In my recent post Mass Extinction in the West Asian Cluster I discussed Eric H. Cline’s book 1177 B.C.: The Year Civilization Collapsed, and in that discussion I characterized the Late Bronze Age (LBA) simultaneous collapse of many civilized societies as a “mass extinction” of civilizations. In the exposition of my argument I first formulated the following idea:
“…civilization in the region likely developed in a kind of reticulate pattern, rather than in a unitary and linear manner, so that, if we were in possession of all the evidence, we might find a series of developments took place in sequence, but not necessarily all originating in a single civilization. Developments were likely distributed across the several different civilizations, and disseminated by idea diffusion until they reached all the others. This could be called a seriation of distributed development.”
This idea, as I now see, can be understood on its own as a distinctive process of complex adaptive systems, applicable not only to civilizations, but also to a range of emergent complexities like life, consciousness, and intelligence as well.
Now I’d like to apply this idea to life, and life under the special circumstances (not presently obtaining within our own planetary system, though that may have been the case in the past) of a multi-planet ecosystem. What, then, is a multi-planet ecosystem?
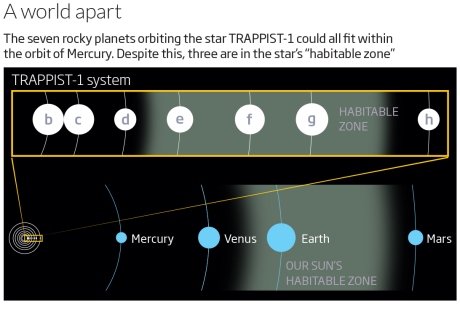
When the TRAPPIST-1 planetary system was discovered, with seven smallish, rocky planets tightly orbiting a small star, the possibilities for life here were of immediate interest to astrobiologists. It has long been thought that lithopanspermia (the transfer of life between planets on rocks) may have occurred within our solar system between Venus, Earth, and Mars — all smallish, rocky planets relatively close in to the sun, and which are known to have have exchanged ejecta from collisions. With an even greater number of small rocky planets in even closer proximity, the likelihood of lithopanspermia at TRAPPIST-1 (assuming life is present in some form) would seem to be higher than in our solar system.
I already know of two papers on the possibilities of lithopanspermia in the TRAPPIST-1 system, Enhanced interplanetary panspermia in the TRAPPIST-1 system by Manasvi Lingam and Abraham Loeb, and Fast litho-panspermia in the habitable zone of the TRAPPIST-1 system, by Sebastiaan Krijt, Timothy J. Bowling, Richard J. Lyons, and Fred J. Ciesla. There is also a paper about the possibilities for botany in the system, Comparative Climates of TRAPPIST-1 planetary system: results from a simple climate-vegetation model by Tommaso Alberti, Vincenzo Carbone, Fabio Lepreti, and Antonio Vecchio.
In a couple of Tumblr posts, More is Different and Yet Another Astrobiology Thought Experiment I discussed some of these possibilities of lithopanspermia in the TRAPPIST-1 system. (And the same interesting TRAPPIST-1 system was also discussed on The Unseen Podcast Episode 69 — A Taste of TRAPPIST-1.)
In More is Different I wrote…
“It may well prove that more is different when it comes to planets, their biospheres, and ecosystems spanning multiple planets. Multi-planet ecologies (we can’t call them biospheres, because they would be constituted by multiple biospheres) may produce qualitatively distinct emergents based on the greater number of components of the ecosystem so constituted. Emergent complexities not possible in a planetary system like our own, with a single liquid-water world, may be possible where there are multiple such planets ecologically coupled through lithopanspermia, and perhaps through other vectors that we cannot now imagine.”
…and in Yet Another Astrobiology Thought Experiment I wrote…
“If life arose separately on several closely spaced planets, with slight biochemical differences between the distinct origin of life events on the several planets, and circumstances within that planetary system were conducive to lithopanspermia, this would mean that each of the planets would eventually have tinctures of life from the other planets, and if these varieties of life could live together without destroying each other, the mixed biospheres of multi-planetary habitable zones where there has been independent origins of life on multiple worlds would suggest a diversity of life not realized on Earth.”
If we combine the ideas of a multi-planetary ecosystem with the idea of reticulate distributed development (which I introduced in relation to civilizational development), we can immediately see the possibility of a multi-planetary ecosystem in which life remains in nearly continuous interaction across several different planets. In such a complex astrobiological context, the great macroevolutionary transitions would not necessarily need to occur all within a single biosphere. It would be sufficient that the macroevolutionary transition took place on at least one planet of the multi-planetary ecosystem, and was subsequently distributed to the other planets of the ecosystem by lithopanspermia. The result would be a seriation of distributed development, i.e., a series of developments taking place in sequence, but not necessarily all originating on a single planet, in a single biosphere. Is this even possible?
We know that microbial life is remarkably resilient, and could likely make the lithopanspermatic journey from one planet to another, but could anything more complex than microbial life make this journey? Recently Caleb A. Scharf in Complex Life: Wimpy or Tough? Complex life may be less resilient than microbial life by some measures, but it’s not necessarily cosmically delicate questions the received wisdom of assuming that eukaryotic multi-cellular life is too vulnerable and delicate to survive “hurdles of selection” — and certainly panspermia must be among the most vertiginous of such hurdles. What about, for example, if conditions were right to freeze complex cells into a still-liquid chamber within a rock, deep in a protected crevice, which then could travel to another planet with complex life intact? There must be similar vectors for panspermia of which we are unaware simply because our imagination fails us.
Obviously, such an occurrence would require many circumstances to occur in just the right order and in just the right way. When this happens for us, as human beings, we say that things are “just right,” and we invoke anthropic selection effects as an explanation, which in this case is simply a Kantian transcendental argument as applied to human beings. But conditions also might be “just right” for some other kind of life, and the antecedent circumstances for such life would be the transcendental conditions of that life — a selection effect of life as we do not know it. This wouldn’t be an “anthropic” explanation in the narrow sense, but if we formalized the concept of an anthropic explanation so that it applied to any being whatsoever, then what human beings call an anthropic explanation would be a special case among a class of explanations. And in this class of explanations would be the “just right” conditions that might lead to rapid and enhanced lithopanspermia among closely spaced planets, which allowed for the transfer to complex life among these planets.
The idea of panspermia has made us familiar with the possibility of life originating on one world and subsequently developing on another world. In case of enhanced and rapid lithopanspermia in an astrobiological context “just right” for such life, we might find life originating on one planet, achieving photosynthesis on another planet, becoming multi-cellular on a third planet, developing an endoskeleton on yet another planet, and so on, possibly continuing to develop into intelligent life. This is what I mean by a seriation of distributed evolutionary development.
If this is possible, if complex life can pass between planets in a multi-planetary ecosystem, I suspect that the rate of evolutionary change would be at least somewhat accelerated in this reticulate astrobiological context, much as the development of civilization was arguably accelerated in the west Asian cluster as a result of the continual interaction of the several civilizations of Mesopotamia, Anatolia, Egypt, and the eastern Mediterranean.
And as life goes, so goes civilization predicated upon life. In a multi-planetary ecosystem, a civilization that grew up on one of these worlds would evolve in a unique astrobiological context that would shape its unique development. Darwin said that, “Man still bears in his bodily frame the indelible stamp of his lowly origin.” Civilizations, too, bear the lowly stamp of their biological origins. A biocentric civilization emergent within a multi-planetary ecosystem would be distinctively shaped by the selection pressures of this ecosystem, which would not be the same as the selection pressures of a single biosphere. And a technocentric civilization arising from a biocentric civilization would continue to carry the lowly stamp of its origins into the farthest reaches of its development.
. . . . .

. . . . .

. . . . .
![]()
. . . . .
![]()
. . . . .


