Civilizations of Planetary Endemism: Part V
4 March 2016
Friday

‘This is an artist’s impression of innumerable Earth-like planets that have yet to be born over the next trillion years in the evolving universe.’
Credits for image and text: NASA, ESA, and G. Bacon (STScI)
Review of Planetary Endemism
So that the reader doesn’t lose the thread of this series on planetary endemism (and to remind myself as well), I began by attempting to formulate a “big picture” taxonomy of planetary civilizations (Part I), but realized that this taxonomy ought to acknowledge the differences in civilization that would follow from civilizations emerging on different kinds of planets (Part II). Then I focused on the question, “What physical gradient is, or would be, correlated with the greatest qualitative gradient in the civilization supervening upon that physical gradient?” (Part III), and next considered how fundamentally different forms of energy flow would beget different kinds of biospheres, which would in turn result in different kinds of civilizations supervening upon these biospheres (Part IV).
This discussion of planetary civilization in terms of planetary endemism provides a new perspective on how we are to understand a civilization that has expanded to the limits dictated by planetary constraints. I have learned that most attempts to discuss planetary civilization get hung up on assumptions of global political and legal unification, which then inevitably gets hung up on utopianism, because nothing like global political and legal unification is on the horizon so this can only be discussed in utopian terms. Thinking about civilization, then, in terms of planetary endemism allows us to get to the substance of planetary civilization without getting distracted by utopian proposals for world government. And what I find to be the substance of planetary civilization is the relationship of a civilization to the intelligent species that produces a civilization, and the relation of an intelligent species to the biosphere from which it emerges.

Thinking about biospheres
How can we scientifically discuss biospheres when we have only the single instance of the terrestrial biosphere as a reference? In order to discuss planetary civilizations scientifically we need to be able to scientifically discuss the biospheres upon which these civilizations supervene. We need a purely formal and general conception of a biosphere not tied to the specifics of the terrestrial biosphere. Ecology is not yet at the stage of development at which it can make this leap to full formalization, but we can make some general remarks about biospheres, continuous with previous observations in this series.
In the Immediately previous post in this series, Part IV, I considered the possibilities of biospheres that fall short of expanding to cover the entire surface of a planet, and so are not quite a biosphere, but constitute what we might call a partial biosphere. In that post I mentioned the terminological difficulties of finding an appropriate word for this and suggested that topology might provide some insight.
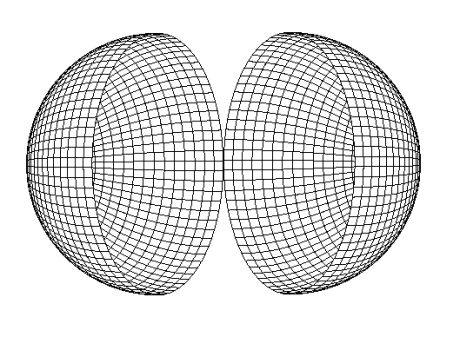
Biospheres and Partial Biospheres
In topology, a biosphere would be what is called a spherical shell, which is bounded by two concentric spheres of different radii. This is the three dimensional extrapolation of what mathematicians call an annulus, which is the area bounded by two concentric circles of different radii. Understanding the biosphere as a spherical shell is a good way to come to an appreciation of the “thickness” of the biosphere. The Terrestrial biosphere may be understood as that spherical shell bounded by the deepest living microbes as the shorter radius and the upper atmosphere as the longer radius. The entry on Deep Subsurface Microbes at MicrobeWiki states: “In oceanic crusts, the temperature of the subsurface increases at a rate of about 15 degrees C per kilometer of depth, giving a maximum livable depth of about 7 kilometers.” The convention establishing the distinction between the upper atmosphere and extraterrestrial space is the Kármán line, 100 km above Earth’s surface. Taking these as the deepest and highest figures, the terrestrial biosphere is a spherical shell approximately 107 km thick, though more conservative numbers could also be employed (as in the illustration above).
A partial biosphere that failed to expand across an entire planetary surface would in topological terms be a punctured spherical shell. Now, a punctured spherical shell is continuously deformable into a sphere, making the two topologically equivalent. This may sound a bit strange, but there is an old joke that a topologist is someone who can’t tell the difference between a doughnut and a coffee cup: each is continually deformable into the other (i.e., both are topologically equivalent to a torus, which is what topologists call a genus 1 surface). In topological terms, then, there is little difference between a biosphere and a partial biosphere (I will discuss a prominent exception in the next installment of this series).
While there is no topological difference between a biosphere and a partial biosphere, there could be a dramatic ecological difference, as a partial biosphere that covered too small of a proportion of a planetary surface would at some point fall below the threshold of viability, while, at the other end of the scale, if it becomes sufficiently extensive it passes the threshold beyond which it can support the evolution of complex life forms. And since only complex life forms produce civilizations, there may be a threshold below which a partial biosphere cannot be associated with a biota of sufficient complexity to allow for the emergence of an intelligent species and hence a civilization.
The extent of a biosphere may place a constraint upon life and civilization emerging from smaller celestial bodies, such as exomoons. So it is not only the possibility of a partial biosphere that may limit the development of complexity in a biota. On the other hand, a system of exomoons, i.e., several inhabitable exomoons orbiting an exoplanet, may have the opposite effect, serving as a speciation pump, leading to higher biodiversity and the emergence of higher forms of emergent complexity. Earlier I suggested that astrobiology is island biogeograpy writ large; a system of inhabitable exomoons, each with its own biosphere, orbiting an exoplanet would offer a particular elegant test of this idea, should we ever discover such a system (and in the immensity of the universe it seems likely that something like this would have happened at least once).
The topology of the biology of a system of exomoons no longer even approximates a biosphere, and this points to the limitation of the concept of a biosphere, and the need for a formalized science of inhabitability that is applicable to any inhabitable region whatever. However, this still is not sufficient for our needs. We must recognize the degree of biological relatedness or difference among separate but biologically related worlds as in the example above.

“This artist’s concept illustrates a quasar, or feeding black hole, similar to APM 08279+5255, where astronomers discovered huge amounts of water vapor. Gas and dust likely form a torus around the central black hole, with clouds of charged gas above and below. X-rays emerge from the very central region, while thermal infrared radiation is emitted by dust throughout most of the torus. While this figure shows the quasar’s torus approximately edge-on, the torus around APM 08279+5255 is likely positioned face-on from our point of view.” (Image and text: NASA/ESA)
The long tail of planetary habitability
However exotic the topology of biospheres to be found in the universe, the biochemistry that populates these biologically connected regions is likely to be constrained by the chemical makeup of the universe. This chemical makeup seems to point to vaguely anthropocentric conditions for life in the universe, but this should not surprise us, as it would be a confirmation of the principle of mediocrity in biology. Water and carbon-based biochemistry is the basis of life on Earth, and the prevalence of these elements in the cosmos at large suggests this as the most common basis of life elsewhere.
Not only are there likely to be liquid subsurface oceans on Europa, Enceladus, and other moons of the outer solar system, possibly with a greater total amount of water on some of these small moons than in all the oceans of Earth, so that we know our solar system possesses enormous resources of water, but we now also know that the universe beyond our solar system possesses significant water resources. The discovery of water vapor at the quasar APM 08279+5255 (described in Astronomers Find Largest, Most Distant Reservoir of Water) represents the presence of vast amounts of water 12 billion light years away — so also 12 billion years in the past — demonstrating both the pervasive spatial and temporal distribution of water in the universe. Astrobiologists have been saying, “To find life, follow the water,” but we now know that following the water would take us far afield.
In additional to water being common in the universe, carbon-based organic chemistry is also known to be common in the universe:
“Astronomers who study the interstellar medium… have found roughly 150 different molecules floating in space… The list boasts many organic (which is to say, carbon-containing) molecules, including some sugars and a still controversial detection of the simplest amino acid, glycine…”
Seth Shostak, Confessions of an Alien Hunter: A Scientist’s Search for Extraterrestrial Intelligence, Washington, DC: National Geographic, 2009, p. 260
Thus, not only is water pervasively present in the universe, but so also are the basic molecules of organic chemistry. I had something like this in mind when in previous post (and elaborated in Not Terraforming, but Something Else…) I tried to outline what might be called variations on the theme of carbon-based life:
“…if life in the outer solar system is to be found, and it is significantly different from life of the inner solar system, how do we recognize it as life? How different is different? It is easy to imagine life that is different in detail from terrestrial life, but, for all intents and purposes, the same thing. What do I mean by this? Think of terrestrial DNA and its base paring of adenine with thymine, and cytosine with guanine: the related but distinct RNA molecule uses uracil instead of thymine for a slightly different biochemistry. Could something like DNA form with G-U-A-C instead of G-T-A-C? Well, if we can consider RNA as being ‘something like’ DNA, then the answer is yes, but beyond that I know too little of biochemistry to elaborate. As several theories of the origins of life on Earth posit the appearance of RNA before DNA, the question becomes whether the ‘RNA world’ of early life on Earth might have also been the origin of life elsewhere, and whether that RNA world matured into something other than the DNA world of terrestrial life.”
I think this is similar to some of the points made by Peter Ward in his book Life as We Do Not Know It, in which Ward wrote:
“…the simplest way to make an alien would be to change DNA slightly. Our familiar DNA is a double helix made up of two on strands of sugar, with the steps of this twisted ladder made up of four different bases. The code is based on triplet sequences, with each triplet either an order to go fetch a specific amino acid or a punctuation mark like ‘stop here.’ Within this elaborate system there are many specific changes that could be made — at least theoretically — that would be ‘alien’ yet might still work.”
Peter Ward, Life as We Do Not Know It: The NASA Search for (and Synthesis of) Alien Life, New York et al.: Penguin, 2005, p. 66-67
Ward considers variations such as changing the backbone of RNA, changing or adding proteins, changing chirality (the direction of the DNA spiral), changing solvents (i.e., a medium for biochemistry other than water), and substituting proteins for nucleic acids. All of these, I think, count as variations on the theme of carbon-based life, which is what we are to expect in the universe rich in carbon-based organic molecules.
Alternative biochemistries with methane-metabolizing microorganisms as described in the recent paper Methane metabolism in the archaeal phylum Bathyarchaeota revealed by genome-centric metagenomics might also be consistent with the dominant chemistry observed in the universe, and would constitute slightly more exotic variations on the theme of carbon-based life. Just as we will have investigated the subsurface oceans of the moons of the outer planets and will know how readily biochemistry emerges in these environments before we even pass the threshold of our own solar system to become an interstellar civilization, so too we will have the opportunity within our own solar system to investigate alternative biochemistries in environments such as Saturn’s moon Titan.
Both water and carbon-based organic chemistry are common in the universe during the Stelliferous Era in the same way that planetary surfaces are common loci of energy flows during the Stelliferous Era; indeed, planetary surfaces provide the vehicle upon which water and carbon-based organic chemistry can produce emergent complexity from energy flows.
The observable universe, then, is rich in planets, in water, and in organic molecules — everything for which one might hope in a search for life. There is no reason for our universe not to be a living universe, in which biochemistry is as common — or will be as common — as as there are planetary surfaces providing energy flows consistent with life as we know it. However, these multitudinous opportunities for life will be constrained by the prevalent organic chemistry of the universe, and this points to variations on the theme of carbon-based like. Other forms of life may exist as outliers, just as biospheres may be driven by energy flows other than insolation, but these will be unusual.

Provisional conclusions
As a provisional conclusion we assert that the same reasoning that leads us to planetary surfaces as the “Goldilocks” zone for energy flows during the Stelliferous Era also leads us to carbon-based life forms employing liquid water as a solvent during the same period of cosmological natural history.
Having thought a bit about the different kind of biospheres that might be possible given different forms of energy flow (Part IV), I have realized that these are probably outliers, and, if we remain focused on civilizations of the Stelliferous Era, insolation of planetary surfaces will be the primary source of energy flows, hence the primary basis of biospheres during the Stelliferous Era, hence the primary basis of civilization up to the point of development when biocentric civilization transitions into technocentric civilization and is no longer exclusively dependent upon a biosphere.
That being said, other sources of energy flow may play a significant role. Radioactive decay has played a significant role in the temperature of Earth (not taking account of radioactive decay, which was not then known, was the reason for Lord Kelvin’s attack on Darwinian time scales). Extrapolating from our own biosphere, we would expect to see a variety of biospheres in which stellar insolation is supplemented by other drivers of energy flow.
Later in the Stelliferous Era, when planetary systems have a greater proportion of heavy elements (due to the process of chemical enrichment), the habitable zone may move further out from parent stars because of the increased availability radioactive decay and natural fission reactors contributing relatively more to the energy flows of biospheres. The increased availability of heavier elements may also eventually impact biochemisty, as forms of life as we do not know it become more likely as the overall mixture of chemicals in the universe matures. The farther we depart in time from the present moment of cosmological natural history, the farther we depart from likely energy flows and biota depending upon these energy flows, until we reach the end of the Stelliferous Era. All that I have written above concerning the Stelliferous Era will cease to be true in the Degenerate Era, when stellar insolation ceases to be a source of energy flows.
For the time being, however, throughout the Stelliferous Era we can count on certain predictable features of life and civilization. Civilization follows intelligence, intelligence follows complex life, and complex life follows from habitability that passes beyond the kind of thresholds described above. Thus the cohort of emergent complexities found in the Stelliferous Era can be traced to the same root.
We may even discover that planetary biospheres exhibit a kind of convergent evolution, not in terms of specific species, but in terms of the kind of biomes and niches available, hence ecological structures to be found, and even the kinds of civilizations supervening upon these ecological structures. For example, I wrote a post on Civilizations of the Tropical Rainforest Biome: on another world with a peer biosphere and an intelligent species, any civilizations we found emergent in the equivalent of a tropical rainforest biome (high temperatures and high rainfall year round) would probably share certain structural features with civilizations of the tropical rainforest biome found on Earth.
The civilizations of planetary endemism, then, include all those classes of sub-planetary civilizations defined by regional biomes, prior to the emergence of a planetary civilization. Each regional (sub-planetary) civilization is consistent with its biome (i.e., it can supply the needs of its agents with the resources available within the biome in question), and in so far as the resources in a given biome govern what is possible for a biocentric civilization emergent within that biome, each such civilization is forced into a kind of uniformity that the institutions of civilization then take up in a spirit of iteration and refinement of a model (i.e., the iterative conception of civilization). When civilization expands until civilizations emergent in distinct biomes are forced into contact, resulting in communication, commerce, and conflict, new forms of planetary scale uniformity emerge in order to facilitate interchanges on a planetary scale.
. . . . .
Planetary Endemism
● Civilizations of Planetary Endemism: Introduction (forthcoming)
● Civilizations of Planetary Endemism: Part I
● Civilizations of Planetary Endemism: Part II
● Civilizations of Planetary Endemism: Part III
● Civilizations of Planetary Endemism: Part IV
● Civilizations of Planetary Endemism: Part V
. . . . .

. . . . .
![]()
. . . . .
![]()
. . . . .


